|
害虫防除に使われる「生物農薬」として登録されている防除資材は、寄生バチや捕食性カメムシなどの昆虫、カブリダニなどの捕食性ダニ、昆虫寄生性あるいは捕食性の線虫、BTをはじめとする昆虫病原微生物、そしてフェロモンなどの生物産生物質の4つに分類されている。
昆虫病原微生物である天敵ウイルスを利用した殺虫剤として、チャのハマキムシ類をターゲットにした顆粒病ウイルス製剤が2003年春に登録され、さらにハスモンヨトウ用の核多角体病ウイルス製剤の登録試験が進行中である。本稿では、天敵ウイルス資材の現状と研究動向について述べたい。
核多角体病ウイルスと顆粒病ウイルス
害虫防除に使われる主要な天敵ウイルス資材は、バキュロウイルス科に属する核多角体病ウイルス(NPV)と顆粒病ウイルス(GV)である。科の名称は、棒状のウイルス粒子の形に由来する。通常、ウイルスの研究には電子顕微鏡による観察や超遠心機を用いた精製が必須であるが、NPVとGVはウイルス粒子を包み込むタンパク質の微小な塊(NPVでは多角体、GVでは顆粒体、総称して封入体)を形成し、NPVの多角体は1~5μm程度(第1図)、GVの顆粒体は長径が300~500nm(第2図)であるため、これらを光学顕微鏡で観察することによって感染の有無が判別できる。また、封入体に包まれた状態のウイルス粒子は比較的安定で、封入体ごと高速遠心機で分離できるなど、他のウイルスに比べて扱いやすい特性を持つ。
バキュロウイルスは経口感染し、封入体で汚染された植物を摂食した幼虫の消化管(中腸)の上皮細胞がウイルス侵入の場となる。チョウ目幼虫の消化液は強アルカリ性であり、消化液で溶かされた封入体から遊離したウイルス粒子が中腸細胞に侵入して増殖を開始する。バキュロウイルスでは、細胞間の感染を担う出芽型粒子と個体間の感染を担う包埋型粒子の2つのタイプのウイルス粒子が形成されるが、天敵ウイルス資材の主成分である多角体と顆粒体に入っている粒子は後者である。
微生物農薬としての天敵ウイルスの現状
微生物農薬として実用化されたウイルスの中で最も施用面積が広いのが、南米で大豆害虫Anticarsia gemmatalisの防除に使われているNPVで、2001/2002年作ではブラジルで155万ha(ダイズ作付面積の11%以上)、パラグアイで約15万haに施用された。
ブラジルにおけるA.gemmatalis NPVの実用化試験は1980年代始めにEnbrapa(ブラジル農牧畜研究公社)で開始され、1990年以降はEnbrapaと契約を結んだ5つの企業に生産、製剤化、品質管理などの技術が移転され、大規模なウイルス生産が行なわれている。
Enbrapaは、品質管理が容易で計画的な生産が可能な実験室内でのウイルス生産方式(人工飼料で飼育した宿主幼虫を用いる)を推奨したが、企業はコストの低い圃場生産方式を選択し、現在ではこれが主体となっている。
圃場生産方式とは、A.gemmatalisが多発生しているダイズ圃場にウイルス散布を行ない、感染時期を見計らって1採集地当り200~300人を動員して幼虫を採集する方法で、日当り採集量は最大で病死虫600kg(30,000haの施用量に相当)に達する。低賃金の労働力が利用できる地域ならではの生産方法である。
ウイルス製剤は、化学農薬に比べ防除コストを20~30%低減することができ、散布回数も少なくてすみ、環境保全のみならず経済面でも有益であることから、近年さらに需要が伸びている。ブラジルではダイズ害虫の種類が限られていること、経済的被害許容水準が高いことなどもウイルス製剤による害虫防除に成功をもたらした要因と考えられる。
この他にもヨーロッパや米国において果樹害虫コドリンガCydia pomonellaのGV、中国やオーストラリア、インドではタバコガ類のNPVなど、さまざまなチョウ目害虫に対してウイルスによる防除が実施されている。各国のウイルス殺虫剤登録状況、入手先などの情報は、無脊椎動物病理学会のホームページ(http://www.sipweb.org/directorymcp/viruses.htm)などで得ることができる。
日本では、過去にハスモンヨトウのNPVやクワゴマダラヒトリのNPVで比較的大規模な実用化試験が行なわれたほか、チャノコカクモンハマキとチャハマキを対象にしたGVの実用化が進められ、鹿児島県で広く普及した。鹿児島県では、県内の茶園面積の約6割でハマキムシ類の防除にGVが利用されている。年1回のGV製剤適期散布で長期間持続する防除効果が得られるため、在来天敵の保護・活用による農薬散布回数の減少が可能となった。
鹿児島県での成功は、防除の機械化、ウイルス製剤の安定生産に加え、農家との連携により適期防除を広域的に実施できる体制が整備されていたことに負うところが大きい。2種類のハマキムシのGVを混合したウイルス製剤は、1990年代始めに国の補助事業によって整備された鹿児島県内の生産施設で、農家の委託による自主生産が行なわれてきたが、農薬取締法の改正に対応する形で2003年3月に農薬登録された。なお、ハスモンヨトウNPVも現在、農薬登録に向けた効果試験が進められている。
ウイルスの感染力向上に関する研究
国内外で行なわれた多くのNPV、GVによるチョウ目害虫防除の研究成果によって、これらのウイルスが化学農薬に偏らない新しい害虫防除体系の一翼を担う素材であることは認識されつつある。しかし、実用化試験を経て、登録、市販に至ったウイルス殺虫剤の数は、世界的に見てもまだ少数であり、効果の安定性向上や生産コストの低減につながる技術の開発は今後の重要課題である。
特に、紫外線防御と感染力強化は、ウイルスの安定性向上や低コスト化実現のポイントであり、活性物質の探索により、様々な化合物やタンパク質が見いだされた。化合物では、スチルベン系蛍光漂白剤が強力な感染促進作用を持つことが米国の研究グループによって明らかにされ、各種害虫とウイルスを用いてその効果が調べられた。
国内でも、アワヨトウなどヤガ科幼虫を用いて、効果の確認ならびに機構の解明が進められ、宿主の消化管を保護している中腸囲食膜の成分であるキチンに蛍光漂白剤が結合して膜形成を阻害するため、ウイルス感染が促進されることが示された。高い感染促進効果が得られ蛍光漂白剤の濃度は1%と高いため散布後の作物が白くなること、農薬としての安全性確認や、環境影響の評価など、実用化に際してはクリアしなければならない課題は多いが、蛍光漂白剤による感染促進の機能解明によって新たな活性物質の探索や開発が進むものと期待される。
この他、生物産生物質として、GVならびに昆虫ポックスウイルス(EPV)由来のタンパク質からもNPV感染促進活性を持つものが見つかっている。これらのNPV感染促進作用は、NPVとGVあるいはNPVとEPVの混合接種実験において発見され、GVとEPVのゲノム上にそれぞれコードされている活性タンパク質遺伝子が特定されている。
感染力の向上によってウイルス感受性の低い宿主を防除対象に加えることが可能になれば、対象害虫の範囲拡大効果も期待できる。筆者らの研究室では、GVのNPV感染促進作用を活用した新たなウイルス製剤の開発を目標に、2004年から5年間のプロジェクトを実施する。
ゲノム解析を中心とした研究の動向
バキュロウイルス研究は、1990年代以降、遺伝子解析を中心に大きな発展を遂げた。
Autographa californica NPV(AcNPV)で初めて全ゲノム塩基配列が報告された1994年以降、現在までに、日本で分離されたカイコNPV、チャノコカクモンハマキNPV、シロモンヤガGV、コナガGVを含む、NPV16種類、GV7種類の全ゲノム塩基配列が遺伝子データベースGenBankで公開された。
この他にも、複数のウイルスでゲノムの部分的な塩基配列情報が公開されており、バキュロウイルスの遺伝子情報の蓄積は、今後さらに加速するものと思われる。遺伝子解析の結果から、バキュロウイルスのゲノムサイズは、100~180kbp、ゲノム上には120~180個の遺伝子が存在すると推定されている。
ウイルスの増殖に適した培養細胞実験系が確立されたAcNPVやカイコNPVは、培養細胞内での相同組換えによって外来遺伝子をウイルスに導入する技術が開発され、組換えバキュロウイルスを使ってタンパク質を発現するシステムがキットで発売されるなど、医学をはじめさまざまな研究分野で利用されている。
1990年代後半には、この組換え技術を利用して、昆虫に特異的な毒素遺伝子を導入したNPVや、特定の遺伝子を欠損あるいは部分的に改変したNPVが作成され、殺虫機能の向上が図られた。欧米では野外試験も一部実施されたが、その後中心となっていた企業が開発研究から撤退し、組換えウイルスの実用化研究は中断された。他方、中国では、現在もオオタバコガNPVを用いて組換えウイルスの開発が行なわれ、野外試験が実施されている。
最近のバキュロウイルス研究は、ゲノム塩基配列情報をもとに特定の遺伝子を欠損あるいは付加したウイルスを用いた遺伝子の機能解明へと進みつつある。これらの成果によって明らかにされたウイルス感染の巧妙なしくみの一端を以下に紹介する。
ウイルスは生きた細胞でしか増殖できないため、宿主側の生物には、植物の過敏感反応のように、ウイルスが侵入した細胞を新たなウイルスが放出される前に自殺(アポトーシス)させるかたちで生体防御するものがあることが知られている。
バキュロウイルスはこれに対抗し自己増殖の場である宿主細胞を維持するため、アポトーシス阻害遺伝子を持っている。アポトーシスは、生物の発生や変態、細胞のガン化などにも深く関わる現象であるため、バキュロウイルスで初めて発見されたアポトーシス阻害遺伝子は医学や理学などの幅広い分野から大きな注目を浴びた。これをきっかけに、その後様々な生物でアポトーシス阻害遺伝子が発見されている。
また、バキュロウイルスの多くは宿主の脱皮ホルモンを不活性化する酵素(エクジステロイドUDP-グルコシルトランスフェラーゼ:EGT)遺伝子を持つことが明らかにされている。この遺伝子は、幼虫脱皮や変態などにともなう幼虫の生理的な変化を制御し、感染幼虫の体をウイルス増殖に適した状態に維持する働きをしているものと考えられる。EGT遺伝子を欠損させたウイルスでは、野生株ウイルスに比べ死亡までの日数が短縮することが明らかにされ、EGT遺伝子は、ウイルスの防除機能向上の面からも注目されている。
この他にも、ウイルスゲノム上にコードされているタンパク質分解酵素(システィンプロテアーゼあるいはカテプシン)とキチン分解酵素に関して興味深い研究成果が得られている。この2つの遺伝子は、ウイルス感染幼虫にみられる、皮膚が裂けやすくなる、死後速やかに体内容が液状化してとろけたようになるなどの現象に深く関与しており、各遺伝子を欠損させた組換えウイルスを感染させた幼虫では、死後数日間死体の形が保たれた。これらの遺伝子は、死体を崩壊させることによってウイルス封入体の拡散を促進する機能を果たしているものと考えられる。
バキュロウイルスが持つ遺伝子の多くはまだ機能が不明のままであり、今後も個々の遺伝子に関する詳細な研究を積み重ねていく必要がある。宿主であるチョウ目昆虫のゲノム解析もカイコやオオタバコガを中心に進展を見せており、その成果はウイルス研究にも寄与するものと思われる。
ウイルスが宿主昆虫を乗っ取り、自己増殖の場を築く仕組みには新たな害虫防除技術の開発につながる知見が数多く含まれている可能性があり、今後のバキュロウイルスの感染機構解明に期待したい。
(独立法人 農業・生物系特定産業技術研究機構 中央農業総合研究センター 虫害防除部)
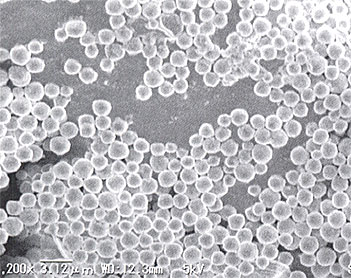
▲第1図 ヨトウガNPV多角体の走査電子顕微鏡像
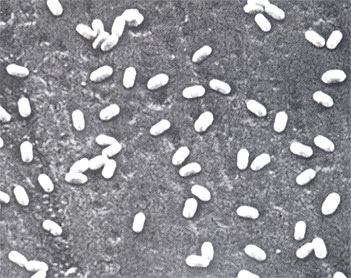
▲第2図 シロモンヤガGV顆粒体の走査電子顕微鏡像
主な参考文献
後藤千枝 (1999).植物防疫 53:303-307.
橋本義文ら (1998).ウイルス 48:51-57.
福原敏彦 (1998).ウイルス 48:59-65.
Hunter-Fujita,F.R.et al.(eds) (1998).Insect Viruses and Pest Management.Chichester:John Wiley & Sons Ltd.
Moscardi,F.(1999).Annual Review of Entomology 44:257-289.
Moscardi,F.et al.(2002).Proceedings of VIII International colloquium on Invertebrate Pathology and Microbial Control.pp.86-91.
仲井まどか、国見裕久 (2000) .微生物の資材化:研究の最前線 鈴井孝仁[ほか] 編、東京、ソフトサイエンス社.pp180-193.
西 八束・野中壽之 (1996).植物防疫 48:469-473.
|
