
|
|
1.はじめに近年、農業生産技術の開発においても、環境調和型農業技術とか永続型農薬技術とか盛んにいわれるようになってきた。病害虫防除技術においても同様である。しかし、わが国における病害虫防除は化学薬剤に依存しているのが実状である。その使用による諸問題の発生にもかからわず、使用を止めるわけにはいかない現実を踏まえ、これからの害虫防除技術体系の確立を目指さねばならない。このような時代背景のもと、微生物殺虫剤の開発研究が活発になり、その利用も多くなってきた。 微生物殺虫剤の素材は天敵微生物である。自然生態系の中で生息している生物は互いに、何らかのかかわり合いをもって生きている。昆虫と微生物とのかかわり合いの中に昆虫とその病原微生物の関係がある。この微生物が天敵微生物ということになる。 微生物殺虫剤で最初に実用化されたのは細菌製剤で、殺虫性芽胞細菌のBacillus thuringiensis(B.t.)とBacillus popilliaeを素材とした製剤が農業生産の現場で使用されているが、BT剤の使用量が圧倒的に多い。
2.BT剤BT剤は1960年、アメリカにおいて商品として出されて以来、世界各国において、害虫防除に使用されている。この素材は前述のようにB.t.菌で、この菌は1911年、ドイツのベルリナーによって、スジコナマダラメイガの病死体から分離され、報告されたが、わが国においては、蚕糸試験場の石渡により、1901年、カイコの卒倒病の病原体として、すでに報告されている。B.t.は通常の土壌細菌で、普通の場所の土壌から容易に分離される芽胞細菌である。 (1)細菌の性状 B.t.の形態は光学顕微鏡観察では、Bacillus cereusと類似しており、識別が困難であるが、両細菌の異なるところは、B.t.は細胞の中に結晶性物体(タンパク質)を産生することである。この結晶性物体が虫を殺すもととなる。 (2)殺虫性毒素 B.t.が産生する殺虫性毒素として殺虫活性が高いものは、デルター内毒素とベーター外毒素である。デルター内毒素は、菌体内に芽胞が形成されるときに、つくられる結晶性物体に含まれているタンパク質で、物体の形は菌株によって特徴がある。写真に示すようなバイピラミッド型・サイコロ型、不定形立方体型などがあり、光学顕微鏡で観察できる。この結晶体は水には溶けないが、還元剤やアルカリ溶液に溶ける。殺虫性タンパク質は亜種(菌株)間で大きさが異なり、24~140kDa(キロダルトン)までの間の複数のタンパク質から成り立っている。その中で殺虫活性を示す部分は分子量5.8~7kDaペプチド断片からなっている。 デルター内毒素産生に関与する遺伝子はプラスミドDNA上か、あるいは染色体DNA上にある。クローニングした殺虫性タンパク質遺伝子の解析が進み、各菌株が産生する殺虫性タンパク質の一次構造(アミノ酸配列)が明らかにされてきた。その結果、各菌株の殺虫性タンパク質構造の共通点と違う点が明らかになり、各菌株をデルター内毒素レベルで分類されるようになった。B.t.の殺虫性タンパク質の遺伝子、分子量、標的昆虫の関係を表に示す。この殺虫性タンパク質遺伝子を他の細菌や植物に導入し、その生物体で発現させることができるようになった。 ベーター外毒素はハエ幼虫に強い殺虫活性を示すことから"fly toxin"ともいわれ、分子量が約700の耐熱性で水溶性のヌクレオチド系の物質である。全てのB.t.菌株がこの毒素を産生するのではなく、一部の菌株のみが産生する。この毒素の活性の作用点がDNA依存RNA合成酵素の阻害にもあるとの報告がだされたことで、多くの国ではこの毒素を産生するB.t.菌株はBT剤として使用が認められていない。
(3)殺虫活性 殺虫性毒素タンパク質を含む結晶性物体は、まず、幼虫のアルカリ性の消化液により溶解され、つづいて、消化液中のタンパク質分解酵素の働きで、部分分解を受ける。その分解物が殺虫活性をもつようになる。この活性化毒素は消化管の中腸上皮細胞に作用する。この毒素の作用を受けた細胞は形が変わり、膨潤し、壊れる。この細胞破壌により、細胞膜による物質の出入ができなくなり、虫は死に至る。以上、デルター内毒素の作用機序を簡単に述べたが、その作用点について、もう少し詳細に説明すると、次のようになる。活性化毒素(ペプチド)は図に示すような構造をもつと考えられている。その毒素は細胞に穴をあける働きをする部分(I)と細胞に結合する部分(II)と、ペプチドの構造安定を保つ部分(III)の三つの部分から成り立っているといわれている。この活性化毒素により、破壌される細胞の細胞膜上には、この毒素と親和性のある特定の場所(レセプター)がある。このレセプターを構成する物質に関する研究が盛んに行なわれている。この物質として報告されているのはアミノペプチダーゼとカドヘリン(細胞間結合物質)である。
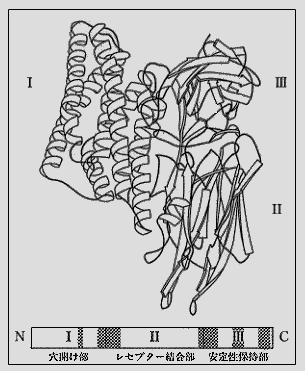
この活性化毒素の作用順序として、現在考えられている説は、この毒素のII部分が細胞膜上にあるレセプターとまず結合する、その結果、毒素の立体構造が変わり、I部分が細胞に穴を開け、細胞を壊す、ということである。 殺虫性毒素タンパク質を含む結晶性物体は昆虫幼虫の消化管を通ることによって、はじめて殺虫活性を示す。この毒素を直接、昆虫の血体腔に入れても、虫は死なない。 デルター内毒素の殺虫スペクトラムは菌株により異なる。鱗翅目昆虫にのみに活性がある菌株、双翅目のみの菌株、鞘翅目だけの菌株、あるいは鱗翅目の中の特定の昆虫にだけ殺虫活性を示す菌株など、多くの特徴がそれぞれの菌株で示される。これらの殺虫活性の違いは、その毒素タンパク質のアミノ酸の配列の違いに基づいている。 昆虫によっては、殺虫活性発現にデルター内毒素と胞子の両者を必要とする場合がある。この胞子の役割は、よく、わかっていないが研究が行なわれている。 (東京農工大学)
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
